Vite

Melo

Mais
Elaborazione risultati in corso
La procedura potrebbe richiedere qualche minuto, si prega di non chiudere il browser fino al completamento dell'operazione.
Al termine dell'elaborazione verrà visualizzata la pagina dei risultati.
Tignola della vite (Eupoecilia ambiguella (Hübner, 1796))
 La Tignola della vite è una specie che predilige climi più umidi e freschi rispetto a L. botrana. Le popolazioni delle due tignole della vite spesso convivono con rapporti quantitativi
influenzati dalle condizioni climatiche locali. La tignola è polifaga e la si può trovare su una trentina di piante appartenenti in particolare ai generi
Vitis, Parthenocissus, Clematis, Cornus, Lonicera, Viburnus, Ligustrum, Ribes, Prunus, Crataegus spp. Tuttavia, le indagini effettuate in Svizzera ed in Friuli-Venezia Giulia
hanno mostrato che la presenza dell'insetto è rara su queste piante selvatiche. La tignola si è rapidamente adattata alla vite coltivata e i suoi danni erano già segnalati nel Medio Evo.
Le uova sono a forma lenticolare con una colorazione bianco-giallastra all'inizio e poi più scure, prima della chiusura. La larva è di colore marrone chiaro da giovane e rosso viola all'ultimo
stadio con il capo nero. L'adulto è una farfalla di circa 10-15 mm con ali anteriori di color giallo biancastro con una fascia nera al centro e le ali posteriori grigio-giallastro.
È un po' più grande della tignoletta e meno mobile. A causa del probabile effetto dell'innalzamento delle temperature, rispetto al passato questa specie sta riducendo la sua diffusione nella regione Friuli Venezia Giulia.
La Tignola della vite è una specie che predilige climi più umidi e freschi rispetto a L. botrana. Le popolazioni delle due tignole della vite spesso convivono con rapporti quantitativi
influenzati dalle condizioni climatiche locali. La tignola è polifaga e la si può trovare su una trentina di piante appartenenti in particolare ai generi
Vitis, Parthenocissus, Clematis, Cornus, Lonicera, Viburnus, Ligustrum, Ribes, Prunus, Crataegus spp. Tuttavia, le indagini effettuate in Svizzera ed in Friuli-Venezia Giulia
hanno mostrato che la presenza dell'insetto è rara su queste piante selvatiche. La tignola si è rapidamente adattata alla vite coltivata e i suoi danni erano già segnalati nel Medio Evo.
Le uova sono a forma lenticolare con una colorazione bianco-giallastra all'inizio e poi più scure, prima della chiusura. La larva è di colore marrone chiaro da giovane e rosso viola all'ultimo
stadio con il capo nero. L'adulto è una farfalla di circa 10-15 mm con ali anteriori di color giallo biancastro con una fascia nera al centro e le ali posteriori grigio-giallastro.
È un po' più grande della tignoletta e meno mobile. A causa del probabile effetto dell'innalzamento delle temperature, rispetto al passato questa specie sta riducendo la sua diffusione nella regione Friuli Venezia Giulia.
Ciclo biologico
La tignola sverna come crisalide, in genere, sotto la corteccia del legno vecchio o in altri anfratti della vite e inizia lo sfarfallamento da inizio aprile in poi; dopo l'accoppiamento, le femmine ovodepongono sui grappolini fiorali. Le larve antofaghe di prima generazione si incrisalidano nel grappolo fiorale ed i nuovi adulti compaiono fra giugno e luglio. Le femmine adulte (a volo crepuscolare) depongono le uova sui bottoni fiorali e sui rachidi. Le larve della seconda generazione sono carpofaghe e hanno un ritmo di accrescimento più lento rispetto alla tignoletta. Lo sfarfallamento della seconda generazione si verifica verso fine giugno - inizio luglio e le farfalle depongono le uova sugli acini in accrescimento. Può verificarsi la presenza di una terza generazione in cui gli adulti depongono le uova sugli acini invaiati, e le larve tendono a rimanere all'interno del grappolo danneggiando alcuni acini.
Sintomatologia e danni
I sintomi e i relativi danni legati alla presenza della Tignola (al pari della Tignoletta) si differenziano (in relazione sia all'entità sia alle parti colpite) in base alla generazione dell'insetto. Le larve della prima generazione attaccano i fiori e successivamente i piccoli acini appena allegati. Gli attacchi delle larve della seconda e terza generazione, invece, sono molto più gravi perché, con la loro attività trofica, attaccano direttamente i grappoli ma favoriscono anche l'attività di funghi e batteri. Ogni larva è in grado di danneggiare da due a più acini contigui. Gli acini colpiti imbruniscono e disseccano, oltre a diventare facile preda di infezioni successive di Botrytis cinerea e/o marciume acido.
Monitoraggio
Per quanto riguarda il monitoraggio di questo lepidottero, si rimanda a quanto riportato nella sezione "Info" del modello per la tignoletta.
Anche per la tignola, le tecniche di monitoraggio sono analoghe a quelle della tignoletta e cioè mediante l'uso di trappole sessuali e/o tecniche di campionamento.
Strategie di difesa
Anche per quanto concerne le strategie di difesa messe in atto per la tignola, esse sono analoghe a quelle descritte per la tignoletta.
Modello previsionale
Il modello fornisce delle stime di numerosità di tutte le fasi fenologiche dell'insetto, permettendo quindi di ridurre i punti di monitoraggio sul territorio con trappole a feromoni e di individuare i momenti ideali per eseguire i monitoraggi in campo. Esso costituisce, inoltre, uno strumento di supporto alle strategie di difesa insetticida.
Risultati elaborazione Tignola
Note sull'utilizzazione: risultati a carattere sperimentale in corso di validazione
Curve fenologiche normalizzate
Fenologia del VITE su scala BBCH sulla base del calcolo dei gradi giorno cumulati (GDD)
| Fase vegetativa | |
| Fase fioritura | |
| Fase accrescimento frutto (solo arboree) | |
| Fase maturazione |
Tignoletta della vite (Lobesia botrana (Denis & Schiffermüller, 1775))
 La tignoletta della vite è un insetto dell'ordine dei Lepidotteri diffuso in tutta Italia, assieme alla tignola è
una delle avversità più pericolose per la coltura della vite rispetto alla quale predilige gli areali viticoli regionali più caldi.
La tignoletta della vite è un insetto dell'ordine dei Lepidotteri diffuso in tutta Italia, assieme alla tignola è
una delle avversità più pericolose per la coltura della vite rispetto alla quale predilige gli areali viticoli regionali più caldi.
Essa attacca principalmente questa coltura, ma è una specie estremamente polifaga, capace di vivere a spese di fiori, foglie e frutti di piante spontanee e coltivate tipiche dell'ambiente mediterraneo anche se non di tutte è conosciuto il ruolo che esercitano su vigore, fecondità, e velocità di sviluppo degli individui. Le larve posso attaccare anche le infiorescenze dell'olivo e possono attaccare anche il kiwi
L'adulto, le cui dimensioni sono circa 10-12 mm di apertura alare, è di colore grigio variegato di giallo-brunastro e di azzurro: la livrea è intensamente marmorizzata. Le larve (9-10 mm di lunghezza) nelle prime fasi di vita sono color ocra-nocciola con il capo scuro, poi il corpo diviene più scuro (verde-brunastro) ed il capo più chiaro.
Ciclo biologico
La tignoletta della vite supera l'inverno allo stadio di crisalide in un piccolo bozzoletto biancastro fra gli anfratti della corteccia della vite, degli eventuali tutori e alla base delle piante. I primi adulti compaiono da aprile a maggio; immediatamente si ha l'accoppiamento e le femmine ovidepongono sui grappolini fiorali. Dalle uova nasceranno le larve della prima generazione, antofaghe (cioè che si cibano dei grappolini fiorali); la loro attività dura circa tre settimane, quindi si incrisalidano nel grappolo originando, all'inizio dell'estate (metà giugno-luglio), gli adulti di secondo volo che ovidepongono sugli acini ben formati, dando origine alle larve di seconda generazione, carpofaghe (cioè che si alimentano del frutto). Queste, dopo la loro attività sugli acini, possono originare le crisalidi svernanti o più frequentemente produrre una terza generazione. In questo ultimo caso lo sfarfallamento degli adulti si avrà da agosto fino a tutto settembre, a seconda degli ambienti. La tignoletta compie, in maniera diffusa, tre generazioni e in certi casi (dovuti all'innalzamento delle temperature medie) si può arrivare anche ad un inizio di quarta generazione.
Sintomatologia e danni
Il danno è determinato dallo stadio larvale. Le larve della prima generazione (generazione antofaga) attaccano i grappolini fiorali e si nutrono dei singoli bottoni fiorali costruendo anche dei nidi sericei, visibili nella parte interna del grappolo; questa generazione non è molto dannosa sia perché il danno ai singoli fiori è sopportabile (per alcune varietà a grappolo serrato potrebbe anche essere un bene), sia perché l'attacco generalmente non è mai massiccio. Le larve della seconda generazione (generazione carpofaga) sono più pericolose perché si nutrono degli acini, entrandovi e svuotandoli. L'attacco è evidente in questo caso, in quanto gli acini avvizziscono ed imbruniscono; ad un esame più attento si notano i fori di entrata o di uscita ed eventualmente anche le larve che si riparano in ricoveri sericei biancastri, costruiti all'interno del grappolo. Le larve della terza generazione si comportano come la precedente, attaccando gli acini e questo avviene a fine estate, quando gli acini sono in fase di maturazione. La seconda e la terza generazione risultano essere le più pericolose in vigneto per le complicanze di natura fungina (botrite e marciume acido) che si possono instaurare sulle ferite provocate dalle larve.
Ricapitolando, si possono generalmente distinguere due tipologie principali di danni causati dalla tignoletta della vite: la prima concerne i danni diretti come la distruzione degli acini, quindi con perdita di prodotto, e dei bottoni fiorali. La seconda consiste in danni indiretti come la diffusione di muffa grigia (botrite) o di marciume acido, con conseguente perdite sia dal punto di vista quantitativo sia dal punto di vista qualitativo del prodotto.
Monitoraggio
Il monitoraggio della tignoletta avviene mediante l'uso di trappole sessuali e/o tecniche di campionamento. Le trappole a feromoni consentono la determinazione dell'inizio dei voli degli adulti e l'evoluzione degli stessi (picco di volo) nelle tre generazioni e quindi permettono di valutare correttamente il posizionamento degli eventuali insetticidi in funzione delle loro caratteristiche (ovicidi, ovolarvicidi, larvicidi). Queste trappole vanno installate (1 o 2 per ettaro o per azienda) a fine marzo; occorre cambiare la capsula con il feromone, ed eventualmente anche il fondo, circa 10 giorni prima del previsto volo della generazione successiva. Il campionamento, invece, si esegue controllando i grappoli (circa 100 ogni ettaro di vigneto), scelti casualmente sul tralcio, su un numero di ceppi randomizzato. Vengono eseguiti in tre epoche prestabilite che corrispondono più o meno alle tre generazioni; in particolare alla fioritura (1a generazione), dalla mignolatura alla pre-chiusura del grappolo (2a generazione) e dalla invaiatura alla prima metà di settembre (3a generazione).
Strategie di difesa
Il metodo migliore per difendersi dalla tignoletta della vite, come per la tignola, è quello della confusione sessuale sia per l'elevata efficacia nel controllo del parassita, sia per il bassissimo impatto ambientale e le nulle interferenze verso la fauna utile del vigneto.
Tale sistema prevede il posizionamento da metà - fine marzo su un ettaro di vigneto di 250-500 diffusori (dispenser-"spaghetti”) che contengono una quantità nota di feromone sessuale della femmina della specie; questo viene rilasciato gradualmente nel corso della stagione, creando una nuvola costante di feromone che non consente più ai maschi di individuare la scia di richiamo rilasciata dalle femmine. In questo modo le femmine non vengono più fecondate e quindi la specie non è più in grado di riprodursi, abbassando notevolmente la presenza di questi lepidotteri nei vigneti. La verifica dell'efficacia del metodo viene condotta sia posizionando all'interno del vigneto "confusionato" 1-2 trappole a feromoni (se queste non catturano maschi, il sistema è efficace), sia con il monitoraggio dei nidi nei grappoli nelle tre generazioni. Il monitoraggio dei nidi larvali nei grappoli risulta essenziale soprattutto nel primo anno di applicazione del metodo perché permette, in ambienti con fortissima pressione dei parassiti, di poter intervenire con eventuali trattamenti insetticidi correttivi nella II e III generazione.
Il metodo si adatta bene a superfici minime di 2 ettari se vigneti isolati oppure anche appezzamenti piccoli se inseriti in comprensori dove tutti i viticoltori applicano la confusione sessuale (ad esempio in Trentino il 100% dei vigneti utilizzano la confusione sessuale o comprensorio Cantina di Casarsa della Delizia o di Ramuscello in Friuli Venezia Giulia).
L'unico inconveniente del metodo è rappresentato dagli erogatori esauriti (dopo due anni di esposizione) che devono essere rimossi dal vigneto in quanto il materiale plastico che li costituisce non è biodegradabile. Le ditte produttrici stanno ultimando gli studi per arrivare all'applicazione di erogatori che siano biodegradabili e quindi rimanere nel vigneto senza ulteriori oneri per il viticoltore. Un metodo che adotta lo stesso principio, ma meno dispendioso per l'azienda dal punto di vista della manodopera nell'applicazione e anche nel ritiro a fine stagione, è l'impiego degli erogatori automatici temporizzati (Buffer) che vengono posizionati in numero di 2.5 – 3 apparecchi per ettaro a seconda delle caratteristiche dei vigneti.
L'altro metodo di lotta è quello che si avvale dell'utilizzo di insetticidi di tipo biologico o di sintesi chimica.
Per le larve della prima generazione, nella maggior parte dei casi, non si prendono in considerazione interventi di tipo insetticida. I trattamenti possono essere effettuati, eventualmente, nella fase dei bottoni fiorali separati, con infestazioni superiori del 30-35% per le varietà da vino con grappolo spargolo e del 50% per le varietà con grappolo serrato. L'intervento insetticida chiave per combattere questo fitofago, qualora non si utilizzi il metodo della confusione sessuale, rimane quello sulla seconda generazione.
Nella scelta dei prodotti chimici da utilizzare è buona norma orientarsi sempre verso quelli a minor impatto ambientale, a minore tossicità per l'uomo e a maggiore selettività.
Le recenti abrogazioni di principi attivi chimici di sintesi hanno modificato in parte le strategie di difesa negli ultimi anni non permettendo interventi insetticidi curativi.
Con insetticidi, regolatori di crescita (IGR) e acceleratori della muta (MAC) è necessario intervenire sulle larve neonate e, in alcuni casi (IGR), sulle uova. Pertanto, il trattamento dovrà essere eseguito a partire dalla fase di ovideposizione che mediamente inizia 3-4 giorni dopo l'inizio delle catture dei maschi nelle trappole a feromoni. La precocità di questo intervento non consente tuttavia di valutare l'effettiva consistenza delle popolazioni e il superamento della soglia di intervento, pertanto si consiglia di riservare questi formulati a vigneti in cui la presenza di tignoletta è particolarmente elevata tutti gli anni.
Un altro gruppo di principi attivi neurotossici ad azione esclusivamente larvicida prevede il posizionamento al momento della fuoriuscita delle prime larve neonate e quindi a circa 7-10 giorni dall'inizio dei voli.
L'unico prodotto biologico attualmente utilizzabile è il Bacillus thuringiensis - var. Kurstaki che agisce per ingestione. Anche per la corretta applicazione di questo insetticida microbiologico è opportuno individuare con precisione il momento della nascita delle larve, in quanto questo prodotto ha una maggiore efficacia all'inizio della loro attività trofica.
Modello previsionale
Il modello fornisce delle stime di numerosità di tutte le fasi fenologiche dell'insetto (ovodeposizione, larve maturate, crisalidi maturate, sfarfallamento adulti) permette di ridurre i punti di monitoraggio sul territorio con trappole a feromoni e di individuare i momenti ideali per eseguire i monitoraggi in campo. Esso costituisce anche uno strumento di supporto alle strategie di difesa insetticida descritte sopra.
Risultati elaborazione Tignoletta
Note sull'utilizzazione: risultati a carattere sperimentale in corso di validazione
Curve fenologiche normalizzate
Fenologia del VITE su scala BBCH sulla base del calcolo dei gradi giorno cumulati (GDD)
| Fase vegetativa | |
| Fase fioritura | |
| Fase accrescimento frutto (solo arboree) | |
| Fase maturazione |
Peronospora della vite (Plasmopara viticola (Berl. & De Toni, 1888))
 La peronospora, causata dal fungo oomicete Plasmopara viticola, è originaria del Nord America e si è diffusa in Europa a partire dalla fine del XIX secolo. Attacca principalmente le varietà di Vitis vinifera, che costituiscono la maggior parte dei vigneti coltivati, e lo sviluppo delle sue epidemie è fortemente influenzato dal clima. È una delle più gravi malattie della vite.
La peronospora, causata dal fungo oomicete Plasmopara viticola, è originaria del Nord America e si è diffusa in Europa a partire dalla fine del XIX secolo. Attacca principalmente le varietà di Vitis vinifera, che costituiscono la maggior parte dei vigneti coltivati, e lo sviluppo delle sue epidemie è fortemente influenzato dal clima. È una delle più gravi malattie della vite.
Ciclo biologico
Il ciclo biologico di Plasmopara viticola include uno stadio sessuato, che garantisce la sopravvivenza dell'agente patogeno in inverno e produce l'inoculo per le infezioni primarie in primavera. Le oospore sono la fase di svernamento del fungo. Lo stadio asessuato è invece responsabile dei cicli di infezione secondaria che si verificano durante la stagione di crescita della vite.
Plasmopara viticola sverna, quindi, sotto forma di oospore nei resti delle foglie colpite dalla malattia.
In presenza di condizioni microclimatiche ben definite, le oospore mature germinano e producono macrosporangi in grado di rilasciare zoospore. In relazione a particolari condizioni di umidità e temperatura si ha l'espulsione di zoospore dai macrosporangi. Le zoospore nuotano nella pellicola d'acqua che bagna la lettiera di foglie; sopravvivono in questa condizione fino a quando l'acqua persiste, ma muoiono rapidamente quando non c'è più acqua. Se durante il periodo di sopravvivenza delle zoospore avviene una pioggia utile, questa produce la dispersione a spruzzo delle zoospore sulle foglie della pianta di vite. La possibilità che le zoospore, arrivate su una foglia recettiva, causino infezione dipende da una combinazione di condizioni favorevoli di temperatura e durata di bagnatura. Le zoospore nuotano, quindi, all'interno dello strato d'acqua presente sulla foglia umida fino ad arrivare agli stomi della pagina inferiore delle foglie, nei quali penetrano. Se l'infezione ha successo, il patogeno si sviluppa all'interno del tessuto fogliare, manifestando i primi sintomi solo a sviluppo completato. L'intervallo di tempo tra la penetrazione del fungo attraverso gli stomi e la comparsa dei sintomi è definito periodo d'incubazione. Il periodo d'incubazione è molto variabile e dipende dalla temperatura e dall'umidità dell'aria. Le infezioni che si originano dalle zoospore provenienti dal terreno sono chiamate "infezioni primarie". Durante la primavera, si possono susseguire diverse infezioni primarie, che continuano fino all'esaurimento dell'inoculo di oospore che sono riuscite a maturare nel periodo invernale.
I sintomi delle infezioni peronosporiche su foglia sono contraddistinti da:
- muffa bianca sul lembo inferiore delle foglie e sui tessuti verdi della pianta dovuta alla produzione di sporangiofori e sporangi;
- decolorazioni giallastre della pagina superiore del lembo fogliare, note con il nome di "macchie d'olio", in corrispondenza della sporulazione sulla pagina inferiore.
Gli sporangi, organi di riproduzione asessuata del fungo, possono essere distribuiti su altra vegetazione anche a notevole distanza grazie alla pioggia o al vento. Come nel ciclo primario, anche in quello secondario gli sporangi rilasciano zoospore che nuotano sullo strato d'acqua, che ricopre la pagina inferiore delle foglie, fino a raggiungere gli stomi. Nella riproduzione secondaria è sufficiente la bagnatura determinata dalla rugiada. I cicli della riproduzione secondaria si susseguono più volte nel corso della stagione con sintomi del tutto uguali a quelle delle infezioni primarie.
La formazione delle oospore, organo sessuato di conservazione del fungo, avviene nei nostri ambienti prevalentemente nel periodo estivo o autunnale; le oospore sono in grado di sopravvivere per svariati anni nei residui fogliari e nel terreno del vigneto.
Sintomatologia e danni
Sulle foglie in accrescimento i primi sintomi sono riscontrabili sulla pagina superiore e consistono in zone decolorate (macchie), che possono arrivare 3 cm e più di diametro, dai contorni relativamente regolari. Sono localizzate preferibilmente lungo i margini e risultano clorotiche giallastre e tipicamente traslucide nel caso della maggior parte dei vitigni a uve bianche e rosse, rossastre in alcuni vitigni ad uve nere. Successivamente, tali aree decolorate possono imbrunirsi e necrotizzare. In condizioni di elevata umidità sulla pagina inferiore, in corrispondenza delle zone decolorate (macchie d'olio), si può osservare la formazione di efflorescenze biancastre rappresentate dalle strutture riproduttive del fungo. Se le foglie sono molto recettive e se l'ambiente è molto favorevole alla malattia, comparsa della efflorescenza biancastra e necrosi possono interessare l'intera pagina fogliare. Gli attacchi gravi di peronospora determinano in genere la caduta anticipata delle foglie (filloptosi).
Nel caso di foglie vecchie (ed in particolar modo dopo l'estate) così come su quelle di vitigni resistenti, le macchie di peronospora risultano piccole, di forma poligonale, delimitate tra le nervature (peronospora a mosaico).
I tralci erbacei vengono attaccati in modo particolare in prossimità dei nodi, generalmente a partire da infezioni sui piccioli fogliari. I sintomi consistono in aree prima idropiche, poi livide e successivamente necrotiche. Spesso si originano spaccature longitudinali in corrispondenza delle quali può svilupparsi l'efflorescenza biancastra costituita dalle sporificazioni di Plasmopara viticola. L'azione del fungo sui tralci erbacei determina ipertrofia dei tessuti con conseguente accrescimento unilaterale del tralcio che si piega andando a formare un uncino o una forma a "S". Le infezioni sui tralci sono molto più rare che sulle foglie.
Sui grappoli l'infezione può avvenire attraverso le infiorescenze o gli acini, oppure attraverso il rachide. Prima dell'allegagione possono comparire sintomi caratterizzati da macchie livide, oltre che da ipertrofie deformanti il peduncolo, raspo e racimoli (forme ad uncino o a "S").
Se le condizioni sono favorevoli allo sviluppo del fungo, e l'attacco è avvenuto precocemente quando stomi e lenticelle sono ancora attivi, i grappoli possono venir ricoperti dalle fruttificazioni del patogeno e quindi seccare.
Dopo l'allegagione, gli attacchi di peronospora determinano sui grappoli due diverse sindromi che vengono comunemente chiamate peronospora palese e peronospora larvata.
La peronospora palese è tipica dei giovani grappoli con raspo e racimoli ancora in gran parte erbacei e acini piccoli. L'infezione di tali organi si manifesta con una colorazione plumbea, successivamente vengono ricoperti dalle fruttificazioni del fungo emesse attraverso gli stomi degli acini.
Sui grappoli più vecchi, l'infezione degli acini determina, per lo più senza evidenziare fruttificazioni fungine (da qui il nome di peronospora larvata), una colorazione brunastra, perdita di turgore e avvizzimento. L'assenza di fruttificazioni del fungo è determinata dal fatto che perdono di funzionalità gli stomi e le lenticelle. La peronospora larvata compare, in genere, quando gli acini hanno raggiunto la fase fenologica di acino delle dimensioni di pisello. La Peronospora determina danni diretti alla produzione (peronospora palese e larvata) e danni indiretti determinati dalla riduzione dell'attività fotosintetica della pianta.
Monitoraggio
Il monitoraggio rappresenta l'attività essenziale del viticoltore per conoscere lo stato sanitario del proprio vigneto riguardo i patogeni-parassiti e quindi stabilire successivamente le corrette strategie di difesa (sulla base dello stato fenologico, tipi di molecole impiegate, gli intervalli tra i trattamenti, etc.) che sono illustrate nel paragrafo successivo.
Questa attività è alla base della difesa integrata delle colture come riportato anche nel Piano di Azione Nazionale per l'uso sostenibile dei prodotti fitosanitari (denominato PAN).
Innanzitutto, il controllo dei vigneti (monitoraggio visivo) deve essere svolto sistematicamente durante tutta la fase di sviluppo vegetativo della vite, quindi dal germogliamento alla fase di fine invaiatura - maturazione, periodo nel quale sia gli organi vegetativi (foglie e germogli) sia i grappoli posso risultare suscettibili alle infezioni del fungo, con sensibilità variabile in funzione della fenologia.
Un monitoraggio adeguato deve prevedere almeno una cadenza settimanale, integrato in periodi di elevata suscettibilità della vegetazione o forte pressione del patogeno (esempio condizioni climatiche favorevoli, piovosità elevate o lunghi periodi di bagnatura, accrescimenti rapidi dei germogli, ecc.) da ulteriori controlli intermedi mirati.
I criteri alla base di un buon monitoraggio dei vigneti sono:
- osservazione di tutte le varietà presenti in un appezzamento. Esiste una diversa sensibilità al patogeno in relazione alla varietà di vite (es. Verduzzo friulano ecc. sensibile agli attacchi su foglia, Merlot estremamente sensibile agli attacchi su grappolo, ecc.);
- maggiore accuratezza nelle zone del vigneto che favoriscono il ristagno di umidità o tempi di bagnatura più prolungati. In collina, ad esempio, si dovrà osservare maggiormente i fondivalle o i versanti esposti a nord, in pianura gli avvallamenti o la vicinanza ai corsi d'acqua o scoline;
- controllo attento delle aree con elevata vigoria dovute, ad esempio, a differenze geomorfologiche del suolo o scelte agronomiche (es. concimazione). L'elevato affastellamento della vegetazione aumenta l'umidità e i tempi di bagnatura delle parti interne della stessa e riduce l'uniformità della distribuzione dei fitofarmaci da parte dei macchinari impiegati;
- visionare attentamente le zone del vigneto in cui i trattamenti degli ultimi 15 giorni potrebbero non essere perfettamente efficaci (es. pioggia intervenuta prima dell'asciugatura completa nel caso di prodotti di copertura oppure evento piovoso prima dell'assorbimento completo nel caso di prodotti citotropici o sistemici);
- sfruttare le varie attività svolte nel vigneto per controllare la vegetazione (trattamenti, altre operazioni colturali sia meccaniche che eventualmente manuali) che pur con minore accuratezza dovuta alla velocità di percorrenza dei filari, consentono di individuare situazioni localizzate anomale (es. problemi alla distribuzione dei fitofarmaci - ostruzione ugelli);
- effettuare i controlli percorrendo a piedi i filari;
- annotare sistematicamente il risultato di tutte le osservazioni riportando la data di controllo. Questo serve in caso di riscontro della malattia a capire i motivi della comparsa dei sintomi.
A causa degli oggettivi problemi di tempistiche nello svolgimento dei monitoraggi, i controlli devono essere effettuati a campione, esaminando un numero rappresentativo di organi vegetativi e grappoli e tenendo in considerazione i criteri sovraesposti; non è pensabile, infatti, riuscire a controllare sistematicamente tutte le viti di un appezzamento in tutte le osservazioni.
Normalmente, per foglie e germogli si sceglie di controllare l'intera vegetazione di un numero di viti distribuite casualmente sia lungo i filari verso i bordi dell'appezzamento sia quelle dei filari centrali. Normalmente è necessario controllare almeno 50-100 viti per vigneto, riservando il numero più alto di piante visionate nei periodi iniziale quando la malattia è meno presente nel vigneto, per essere in grado di individuare eventuali focolai localizzati.
Per quanto riguarda i grappoli devono essere osservati almeno 100 per vigneto sempre nelle stesse piante scelte per il controllo delle foglie. La posizione dei grappoli sulla singola pianta deve essere scelta in funzione anche della forma di allevamento; per esempio, nel Sylvoz o Casarsa scegliere un eguale numero di grappoli posti nella parte bassa e quelli della zona del cordone.
In affiancamento alle informazioni ricavate dai monitoraggi aziendali si possono ottenere utili indicazioni generali o territoriali consultando i bollettini settimanali pubblicati da ERSA e dai Consorzi DOC sulla piattaforma di AgriCS nella sezione "Bollettini" o sul sito dell'ERSA.
Strategie di difesa
Per impostare una corretta strategia di difesa bisogna considerare la biologia del fungo, le caratteristiche climatiche dell'area di coltivazione e lo sviluppo fenologico della vite.
Il metodo più noto e diffuso per segnalare l'avvio delle infezioni primarie, e di conseguenza l'inizio dei trattamenti, pur con suoi limiti, è la regola dei "3 dieci":
- superamento della temperatura minima di 10°C, al disotto di questa temperatura si arresta lo sviluppo del fungo;
- lunghezza dei germogli di circa 10 cm, cioè quando, nella maggior parte delle varietà, le foglie diventano recettive (stomi attivi). Questo stadio corrisponde alla dimensione fogliare superiore ai 3 centimetri;
- quantità di pioggia caduta nelle ultime 24/48 ore di 10 mm, necessaria sia a far germinare le oospore che a veicolare le zoospore sulle foglie.
Negli ultimi anni, con la diffusione degli strumenti informatici e di monitoraggio ambientale, sono stati elaborati modelli previsionali accurati ed attendibili che possono essere utilizzati quali validi supporti alle decisioni per la difesa antiperonosporica.
Viste le condizioni climatiche estremamente favorevoli allo sviluppo della peronospora nei periodi primaverile ed estivo, in gran parte della nostra regione, non è pensabile la coltivazione della vite senza una difesa accurata e costante durante tutto il periodo di accrescimento vegetativo della pianta.
Le caratteristiche e i meccanismi d'azione dei principi attivi utilizzabili per la difesa della coltura si possono raggruppare in:
- prodotti contatticidi/copertura che non penetrano nella vegetazione e esplicano la loro azione solo sulla superficie. La resistenza al dilavamento dipende dalle caratteristiche dei formulati e dalla capacità di legarsi alle cere cuticolari;
- citotropici e citotropici traslaminari, che penetrano nel tessuto vegetale senza entrare nella circolazione linfatica. Questa caratteristica li pone al riparo dal dilavamento;
- sistemici, sono prodotti che penetrano nella vegetazione e hanno la caratteristica di spostarsi anche negli organi non trattati, più o meno intensamente per via linfatica;
- prodotti ad azione preventiva, se presenti sulla vegetazione impediscono l'instaurarsi di nuove infezioni;
- prodotti ad azione curativa che sono in grado di interrompere lo sviluppo del fungo specie nelle prime fasi dell'incubazione;
- prodotti ad azione eradicante/antisporulante che impediscono la formazione dei rami sporangiofori e devitalizzano le zoospore;
- prodotti ad azione multisito che agiscono in più processi biochimici del fungo. Questi prodotti sono meno soggetti a selezionare ceppi del patogeno resistenti;
- prodotti ad azione monosito che agiscono solo su uno specifico processo biochimico del fungo. Questi prodotti sono più soggetti a selezionare ceppi del patogeno resistenti.
La strategia di difesa adottata nei nostri areali viticoli prevede:
- il posizionamento preventivo dei prodotti nei trattamenti in relazione alle possibili infezioni prima delle piogge infettanti.
- utilizzo sempre di prodotti di copertura ad azione multisito, all'inizio e a fine stagione da soli, e, dalla fase di prefioritura sino alla chiusura del grappolo, in miscela con prodotti citotropici, endoterapici o affini alle cere.
Al fine di evitare l'insorgenza delle resistenze, i prodotti monosito vengono sempre consigliati in abbinamento con altri principi attivi (possibilmente multisito).
Per quanto concerne la difesa in viticoltura biologica, l'attuale normativa (Reg. CE 834/07) impone forti restrizioni all'uso dei principi attivi impiegabili contro la peronospora della vite. Fra le sostanze consentite, molte hanno dimostrato, nei test sperimentali, una certa efficacia, ma non sempre tale efficacia è riuscita a garantire una protezione affidabile in campo. Attualmente, il principio attivo su cui si basa la difesa in viticoltura biologica è il rame che, purtroppo, presenta forti limitazioni sui quantitativi impiegabili.
Negli ultimi anni si stanno provando nuovi prodotti di origine biologica (lieviti, batteri, estratti vegetali) che integrano la difesa o con azione diretta contro il patogeno o mediante l'attivazione dei meccanismi di resistenza endogeni della pianta.
Modello previsionale
Il modello considera l'evoluzione temporale delle infezioni primarie nelle diverse fasi di sviluppo del patogeno, fornendo i seguenti output: date di germinazione delle oospore del terreno (con formazione degli sporangi), date di rilascio delle zoospore dagli sporangi, date di dispersione delle zoospore sulle foglie e sui tralci delle viti, date di infezione dei tessuti fogliari da parte delle zoospore, periodo di incubazione (periodo di formazione delle macchie d'olio sulle foglie). Fornisce, inoltre, informazioni sulle date più probabili di comparsa delle infezioni secondarie.
Risultati elaborazione Peronospora
Note sull'utilizzazione: risultati a carattere sperimentale in corso di validazione
Infezioni primarie
Infezioni secondarie
Fenologia del VITE su scala BBCH sulla base del calcolo dei gradi giorno cumulati (GDD)
| Fase vegetativa | |
| Fase fioritura | |
| Fase accrescimento frutto (solo arboree) | |
| Fase maturazione |
Cicalina della flavescenza dorata (Scaphoideus titanus (Ball, 1932))
 Insetto appartenente all'ordine Hemiptera (sottordine Cicadomorpha, famiglia Cicadellidae) è una specie originaria del Nord America ed è considerato il vettore principale della Flavescenza dorata.
Le uova di Scaphoideus titanus sono fusiformi e di colore bianco-crema, lunghe circa 1,5 mm. Questa specie presenta cinque stadi giovanili: i primi due vengono chiamati neanidi in quanto sono privi di abbozzi alari. Gli ultimi tre, chiamati ninfe, presentano gli abbozzi alari.
L'adulto è lungo 5,5-6 mm e ha una colorazione di base a bande ocra-brune sulla sommità del capo e sul torace. Nel capo, sono presenti linee nere trasversali e le ali sono di colore bruno con alcune porzioni biancastre, con nervature nere. Le zampe sono di color crema, mentre quelle metatoraciche presentano bande più scure e delle spinette mobili.
Gli stadi giovanili di questo insetto possono essere confusi con altre specie, ad esempio la cicalina verde della vite (Empoasca vitis Göethe, 1875), la cicalina gialla della vite (Zygina rhamni Ferrari, 1882) e la metcalfa (Metcalfa pruinosa Say, 1830). Un tratto distintivo è la presenza sull'ultimo segmento dell'addome di Scaphoideus titanus di due tacche nere simmetriche. Inoltre, a differenza delle cicaline i giovani di Scaphoideus titanus saltano se disturbati, mentre quelli delle cicaline si spostano rapidamente (la verde) o lentamente (la gialla). Metcalfa è distinguibile per l'addome più tozzo e la presenza di secrezioni cerose.
Insetto appartenente all'ordine Hemiptera (sottordine Cicadomorpha, famiglia Cicadellidae) è una specie originaria del Nord America ed è considerato il vettore principale della Flavescenza dorata.
Le uova di Scaphoideus titanus sono fusiformi e di colore bianco-crema, lunghe circa 1,5 mm. Questa specie presenta cinque stadi giovanili: i primi due vengono chiamati neanidi in quanto sono privi di abbozzi alari. Gli ultimi tre, chiamati ninfe, presentano gli abbozzi alari.
L'adulto è lungo 5,5-6 mm e ha una colorazione di base a bande ocra-brune sulla sommità del capo e sul torace. Nel capo, sono presenti linee nere trasversali e le ali sono di colore bruno con alcune porzioni biancastre, con nervature nere. Le zampe sono di color crema, mentre quelle metatoraciche presentano bande più scure e delle spinette mobili.
Gli stadi giovanili di questo insetto possono essere confusi con altre specie, ad esempio la cicalina verde della vite (Empoasca vitis Göethe, 1875), la cicalina gialla della vite (Zygina rhamni Ferrari, 1882) e la metcalfa (Metcalfa pruinosa Say, 1830). Un tratto distintivo è la presenza sull'ultimo segmento dell'addome di Scaphoideus titanus di due tacche nere simmetriche. Inoltre, a differenza delle cicaline i giovani di Scaphoideus titanus saltano se disturbati, mentre quelli delle cicaline si spostano rapidamente (la verde) o lentamente (la gialla). Metcalfa è distinguibile per l'addome più tozzo e la presenza di secrezioni cerose.
Ciclo biologico
Scaphoideus titanus compie una sola generazione all'anno; lo svernamento avviene allo stadio di uovo, deposto sotto la corteccia della vite (solitamente questa specie preferisce il legno di due anni ma anche quello più vecchio, raramente depone le uova sul legno di un anno).
La temperatura è il fattore maggiormente condizionante la dinamica della schiusa, che è rapida in caso di inverni rigidi. Proprio questo fattore sembra avere un ruolo nella regolazione della sincronia tra le schiuse e il germogliamento della vite, che consente alle neanidi di reperire il cibo necessario per la propria sopravvivenza.
In vigneto, le prime neanidi compaiono, di norma, tra la prima e la seconda decade di maggio, mentre gli adulti sono presenti dall'inizio di luglio. Il picco dei voli avviene tra la fine di luglio e la metà di agosto, mentre gli ultimi adulti sono osservabili alla fine di ottobre (e questo è dovuto alle temperature autunnali sempre più miti).
La specie in questione è strettamente ampelofaga: il ciclo biologico può essere cioè completato solo su specie del genere Vitis. I giovani, infatti, sono attratti da sostanze volatili emesse dalle foglie della vite.
Sintomatologia e danni
Gli stadi giovanili determinano la suzione della linfa dalle nervature secondarie e terziarie delle foglie di vite, mentre le ninfe e gli adulti succhiano la linfa dalle nervature principali. È da considerare che l'attività di suzione può avvenire, talvolta, anche a carico dei piccioli fogliari e dei tralci erbacei. Lo Scaphoideus titanus, attraverso la sua attività trofica (di tipo floematico e a volte anche xilematico), determina danni diretti trascurabili e non apprezzabili (considerando che, generalmente, nel vigneto l'insetto è presente con bassa densità di popolazione). Necessitano di una considerazione particolare i danni indiretti, che questa cicalina è in grado di compiere: è, infatti, il vettore del fitoplasma della Flavescenza dorata. Per limitare la diffusione della malattia in questione, data la sua estrema gravità e importanza per le produzioni vitivinicole, nel 2000 è stato emanato un decreto ministeriale di lotta obbligatoria contro lo Scaphoideus titanus e la Flavescenza dorata.
Monitoraggio
Il monitoraggio è necessario per valutare la presenza di Scaphoideus titanus nei vigneti.
La presenza degli stadi giovanili va valutata attraverso controlli visivi in campo. I conteggi vengono effettuati scegliendo casualmente delle piante ed esaminando la pagina inferiore di 5 foglie per pianta, scelte fra quelle (foglie basali e/o sui succhioni) più vicine al legno di due o più anni, dove vengono deposte le uova. In alternativa, la verifica può essere eseguita in laboratorio prelevando i germogli e inserendoli in buste di plastica. I controlli devono essere realizzati nelle prime ore del mattino, non dopo le piogge. Altri metodi di monitoraggio prevedono la cattura mediante il retino entomologico oppure lo scuotimento dei tralci e successiva raccolta degli insetti mediante un telo o un ombrello entomologico.
La valutazione dell'andamento fenologico, eseguita a livello di comprensorio, prevede la cattura degli individui e la successiva determinazione dello stadio giovanile in laboratorio.
Per il monitoraggio degli adulti, generalmente, vengono utilizzate trappole cromotropiche gialle. Per valutare la densità nell'appezzamento queste vanno posizionate nel periodo di maggior presenza, cioè fine luglio – inizio agosto.
Strategie di difesa
Visto che Scaphoideus titanus è strettamente associato alla Flavescenza dorata, la quale è considerata una malattia da quarantena, tale insetto è un organismo soggetto a lotta obbligatoria. Fra le disposizioni obbligatorie c'è il controllo delle popolazioni di Scaphoideus titanus con trattamenti insetticidi; essi vengono eseguiti uno o due volte, in genere tra la fine di giugno e i primi giorni di luglio. La difesa insetticida diventa particolarmente importante dove è presente un'elevata carica di inoculo, costituita dalle piante infette.
Un'altra situazione dove è opportuno effettuare un'efficiente lotta insetticida al vettore si ha nel vivaismo viticolo. In questo caso l'eliminazione preventiva delle popolazioni di Scaphoideus titanus deve avvenire anche in assenza di piante sintomatiche. La vigente normativa impone ai vivaisti una serie di misure molto restrittive, la cui applicazione garantisce la qualità dei materiali assoggettati a certificazione.
Fra i principi attivi utilizzabili, vi sono neonicotinoidi e butenolidi, nonché i piretroidi. In agricoltura biologica è ammesso l'impiego del piretro naturale; altre sostanze attive autorizzate sono i sali potassici di acidi grassi o l'azadiractina.
Per quanto riguarda, infine, le ricerche in corso su mezzi di lotta alternativi, vi sono l'impiego del caolino (argilla inerte, bianca, che distribuita sulle piante le rende meno visibili da parte degli insetti), le reti escludi-insetto, la confusione sessuale vibrazionale, la termoterapia in acqua calda (che sembrerebbe devitalizzare le uova) e il controllo simbiotico (strategia che utilizza la conoscenza delle relazioni tra insetti e simbionti).
Modello previsionale
Il trattamento deve coincidere con determinate fasi fenologiche dell'insetto, per poter metter in atto in maniera corretta i trattamenti stessi ed in accordo con quanto previsto dalla Direttiva 2009/128/CE sull'uso sostenibile dei fitofarmaci. Questo modello, così come quello per la tignoletta e per la carpocapsa, consente quindi di visualizzare l'andamento delle fasi fenologiche dell'insetto (schiusura uova, evoluzione stadi giovanili, neanidi e ninfe, sfarfallamento, adulti) che dipende dall'andamento della temperatura, permettendo così una programmazione degli interventi in campo quanto più efficace e sostenibile e una coretta gestione fitoiatrica di campo.
Risultati elaborazione Scafoideo
Note sull'utilizzazione: risultati a carattere sperimentale in corso di validazione
Fenologia del VITE su scala BBCH sulla base del calcolo dei gradi giorno cumulati (GDD)
| Fase vegetativa | |
| Fase fioritura | |
| Fase accrescimento frutto (solo arboree) | |
| Fase maturazione |
Ticchiolatura del melo (Venturia inaequalis - Fusicladium dendriticum)
La Ticchiolatura rappresenta la più grave e diffusa avversità crittogamica del melo, in grado di causare danni rilevanti se non vengono attuate idonee misure di controllo: questo fungo colpisce foglie, germogli e frutti, sia sulla pianta che in magazzino. Il fungo responsabile della ticchiolatura vive esclusivamente allo stato parassitario e compie il suo ciclo di sviluppo alternando una forma sessuata (Venturia inaequalis) ad una asessuata (Spilocea pomi = Fusicladium dendriticum). La prima si differenzia all'interno dei tessuti dell'ospite quando le condizioni ambientali diventano sfavorevoli e consente al patogeno di perpetuarsi durante i mesi invernali. È caratterizzata da pseudoteci globosi nei quali si formano aschi cilindrici contenenti ciascuno otto ascospore.
La fase asessuata o stadio conidico compare in primavera e ad essa è legata la diffusione nell'ambiente del fungo: le spore agamiche (conidi) si formano all'apice di rami conidiofori che erompono dall'epidermide dei tessuti infetti della pianta.
Ciclo biologico
La Ticchiolatura supera la stagione sfavorevole invernale sotto forma di pseudoteci che si sono formati in autunno, sulla vegetazione colpita (in particolare sulle foglie) caduta a terra. Il fungo, raramente, può svernare come micelio negli anfratti gemmari o sulle tacche rameali. Questa modalità di svernamento è stata osservata solo in piante isolate e molto vigorose.
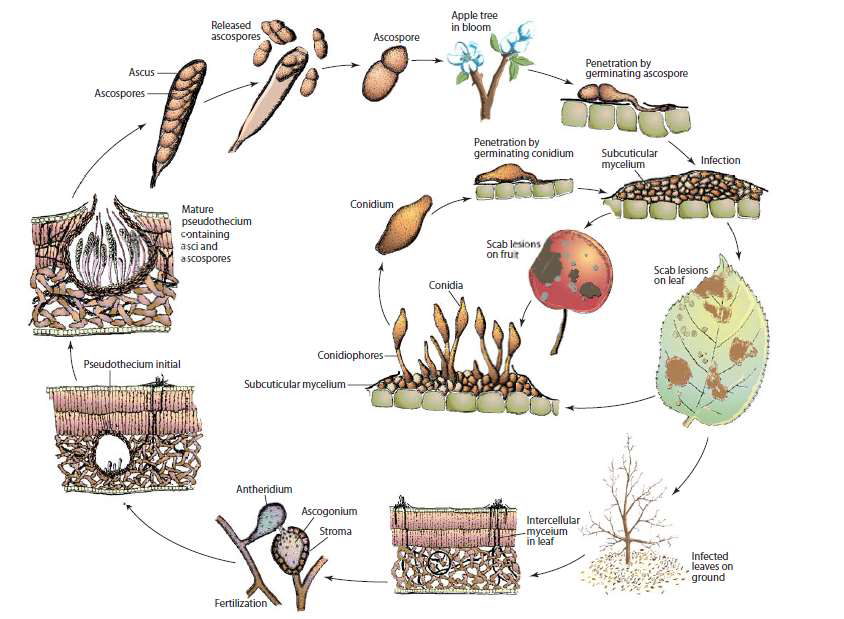 In primavera (seconda-terza decade marzo) con temperature superiori ai 5°C e con le piogge stagionali (sono sufficienti solo 0,2 mm di pioggia), gli pseudoteci di Venturia inaequalis "maturano" e liberano le ascospore che vengono trasportate dal vento o dagli schizzi d'acqua, arrivando sulla vegetazione giovane dove, in condizioni favorevoli, danno origine ad una infezione primaria. Normalmente questa fase dura in media 40-45 giorni, periodo che inizia a fine marzo fino a metà-fino maggio a seconda della stagione. Una volta che il fungo è penetrato nelle foglie, si origina il micelio che si sviluppa tra la cuticola e la parete cellulare delle cellule epidermiche. Dopo un periodo d'incubazione, sugli organi colpiti emerge all'esterno mostrando i tipici sintomi. In presenza di bagnature, dai tessuti colpiti si originano delle fruttificazioni agamiche (conidi) da cui prendono il via le infezioni secondarie responsabili dell'ulteriore diffusione della malattia che si prolungheranno fino alla fine della stagione.
In primavera (seconda-terza decade marzo) con temperature superiori ai 5°C e con le piogge stagionali (sono sufficienti solo 0,2 mm di pioggia), gli pseudoteci di Venturia inaequalis "maturano" e liberano le ascospore che vengono trasportate dal vento o dagli schizzi d'acqua, arrivando sulla vegetazione giovane dove, in condizioni favorevoli, danno origine ad una infezione primaria. Normalmente questa fase dura in media 40-45 giorni, periodo che inizia a fine marzo fino a metà-fino maggio a seconda della stagione. Una volta che il fungo è penetrato nelle foglie, si origina il micelio che si sviluppa tra la cuticola e la parete cellulare delle cellule epidermiche. Dopo un periodo d'incubazione, sugli organi colpiti emerge all'esterno mostrando i tipici sintomi. In presenza di bagnature, dai tessuti colpiti si originano delle fruttificazioni agamiche (conidi) da cui prendono il via le infezioni secondarie responsabili dell'ulteriore diffusione della malattia che si prolungheranno fino alla fine della stagione.
In autunno il ciclo stagionale del fungo si concluderà nelle foglie cadute a terra (fase saprofitaria) con la formazione del micelio secondario che, per cariogamia, formerà i pseudoteci. Questi si conserveranno durante la stagione invernale e daranno origine, nella primavera successiva, alla carica di inoculo primario.
Infezioni primarie:
La quantità di ascospore vitali in un determinato momento della stagione vegetativa (potenziale di inoculo) dipende dal livello di attacco presente nel frutteto a fine stagione e dalle condizioni climatiche che si susseguono fino alla ripresa vegetativa. La quantificazione del potenziale di inoculo, ossia del potenziale infettivo di ogni frutteto responsabile della gravità delle possibili infezioni primarie è tuttora in dibattuta. Tale potenziale, durante la stagione invernale, subisce inevitabilmente una riduzione a causa della decomposizione delle foglie cadute e di un processo di maturazione in prossimità della ripresa vegetativa delle piante.
Nel 1986 McHardy (USA) ha elaborato un sistema di calcolo per stimare il potenziale di inoculo del patogeno o PAD (Potential Ascospore Dose) alla ripresa vegetativa in funzione di due variabili (il n° di macchie su 600 germogli e la percentuale di foglie con ticchiolate) calcolate in seguito a rilievi eseguiti in campo in autunno poco prima della caduta delle foglie. Tuttavia, il suo utilizzo pratico risulta di difficile attuazione e lo stesso autore ha proposto nel 1999 una semplificazione di tale stima basata unicamente sul n° di foglie con ticchiolatura rilevato su un campione di 600 germogli.
Il primavera il volo delle ascospore dipende da diversi fattori, i più importanti dei quali sono:
- Luce: condiziona molto il volo, il 95% delle ascospore viene rilasciata durante il giorno;
- Pioggia: provoca il volo delle ascospore che dal terreno si spostano alla pianta e tale movimento può avvenire con precipitazioni di soli 0,2 mm. La gravità dell'infezione è provocata dalla quantità di spore mature al momento della pioggia e da quante di esse riescono successivamente a germinare;
- Temperatura: determina la maturazione delle ascospore. Piogge successive a periodi con temperature miti comportano voli intensi e rischi elevati di infezioni.
Quindi, in sintesi, la luce e la pioggia sono i fattori in grado di influenzare l'emissione delle ascospore, che associati alla durata della bagnatura fogliare, ad una temperatura ottimale, alla presenza di vegetazione recettiva ed al potenziale di inoculo (quantità di spore mature) sono i fattori che permettono alle spore mature di raggiungere le piante, germinare e dare origine alle infezioni primarie di ticchiolatura di gravità più o meno elevate a seconda del numero delle ascospore evase.
Sintomatologia e danni
Sintomi sulle foglie
 Le prime infezioni sono generalmente visibili sulla pagina superiore, in forma di macchie inizialmente decolorate, distribuite irregolarmente; in seguito, queste lesioni assumono una colorazione più scura e contorni meglio definiti e sono osservabili anche sulla pagina inferiore. Se c'è elevata umidità, i tessuti colpiti si ricoprono di fruttificazioni fungine che conferiscono alle lesioni un aspetto vellutato. Le foglie ticchiolate disseccano e cadono prematuramente, lasciando le piante parzialmente spoglie.
Le prime infezioni sono generalmente visibili sulla pagina superiore, in forma di macchie inizialmente decolorate, distribuite irregolarmente; in seguito, queste lesioni assumono una colorazione più scura e contorni meglio definiti e sono osservabili anche sulla pagina inferiore. Se c'è elevata umidità, i tessuti colpiti si ricoprono di fruttificazioni fungine che conferiscono alle lesioni un aspetto vellutato. Le foglie ticchiolate disseccano e cadono prematuramente, lasciando le piante parzialmente spoglie.
Sintomi sui germogli
Sono simili a quelli sulle foglie; si può osservare una "tacca" cancerosa e persistente anche dopo la lignificazione.
Sintomi sui frutti
 L'attacco sui fiori e sui rametti è in genere meno frequente: si manifesta sotto forma di lesioni brunastre a carico di petali, calice, peduncolo fiorale e, nel caso dei rami, dei tessuti ancora allo stato erbaceo.
L'attacco sui fiori e sui rametti è in genere meno frequente: si manifesta sotto forma di lesioni brunastre a carico di petali, calice, peduncolo fiorale e, nel caso dei rami, dei tessuti ancora allo stato erbaceo.
Monitoraggio
 Per il monitoraggio del volo delle ascospore si utilizzano alcuni strumenti detti captaspore apparecchi elettronici o meccanici che sono in grado di catturare nell'ambiente aereo le ascospore aerodiffuse e di convogliarle su vetrini o nastri di plastica trattati con silicone, per farli adeguatamente aderire. I supporti vengono poi analizzati al microscopio per conteggiare la quantità di spore ed avere indicazioni sul potenziale di inoculo. Il numero di ascospore conteggiate permette di conoscere la gravità di infezione.
Per il monitoraggio del volo delle ascospore si utilizzano alcuni strumenti detti captaspore apparecchi elettronici o meccanici che sono in grado di catturare nell'ambiente aereo le ascospore aerodiffuse e di convogliarle su vetrini o nastri di plastica trattati con silicone, per farli adeguatamente aderire. I supporti vengono poi analizzati al microscopio per conteggiare la quantità di spore ed avere indicazioni sul potenziale di inoculo. Il numero di ascospore conteggiate permette di conoscere la gravità di infezione.
Strategie di difesa
Le tecniche agronomiche di controllo della malattia prevedono l'eliminazione delle foglie infette cadute a terra e la somministrazione di concime a base di azoto (urea) in grado di favorire l'attività microbica del suolo e quindi la degradazione dei residui colturali. Questo ha lo scopo di ridurre l'inoculo per la stagione successiva.
I trattamenti prendono avvio alla ripresa vegetativa del melo nella fase fenologica di "punte verdi" e, se nel frutteto non vengono rilevati sintomi della malattia una volta terminato il periodo delle infezioni primarie, dopo la fase di "frutto noce", quando i frutti sono meno sensibili alle infezioni è possibile adottare turni di interventi più lunghi.
Per gestire al meglio la difesa contro questo fungo nell'areale frutticolo regionale si è visto, per esperienza diretta dei frutticoltori, che è fondamentale controllare le infezioni primarie da subito e cioè dal primo volo delle ascospore. Questo garantisce una maggior tranquillità nella gestione della difesa estiva del frutteto, con una conseguente riduzione degli interventi specifici nel corso dell'estate.
Le attuali strategie di difesa sono basate principalmente su interventi di tipo preventivo mirato, posizionando il trattamento con prodotti di copertura il più vicino possibile ad un evento piovoso, tenendo in considerazione la crescita vegetativa e lo stadio fenologico delle piante, limitando in questo modo l'utilizzo di prodotti ad azione curativa.
Per impiegare correttamente le sostanze attive (S.A.) disponibili è possibile individuare quattro periodi di riferimento:
- La prefioritura ovvero tutto il periodo che intercorre dalla ripresa vegetativa (punte verdi) fino a bottoni rosa;
- La fioritura, che rappresenta la fase di maggior criticità;
- Da allegagione a frutto noce;
- Dopo la fase di frutto noce oltre la quale la recettività dei frutti al fungo diminuisce e con essa la pericolosità della malattia.
Dopo il trattamento, la vegetazione risulterà protetta da 3 a 5 giorni, a seconda del tipo di prodotto utilizzato e della velocità di crescita dei tessuti vegetali. Nel caso di interventi con infezioni in atto, è opportuno eseguire trattamenti a turno ravvicinato con prodotti endoterapici.
Terminato il periodo delle infezioni primarie verificato sia con il monitoraggio in campo del volo delle ascospore sia con l'ausilio dei modelli sperimentali, coincide negli areali di pianura con l'ultima decade di maggio e in montagna con la prima di giugno (a seconda della stagione), la difesa estiva viene pianificata in base alla presenza o meno di infezioni di ticchiolatura in campo, ricorrendo a prodotti preventivi.
Modello previsionale
Il corretto momento per iniziare i trattamenti per la difesa da ticchiolatura viene determinato attraverso l'uso di modelli previsionali, i quali sono in grado di simulare il processo di maturazione degli pseudoteci e di segnalare quando questi giungono a maturazione e, in seguito alle piogge, in grado di rilasciare le ascospore.
Alcuni modelli previsionali della ticchiolatura come RimPro ed Omnia FVG stimano già, in maniera preventiva, l'evoluzione dell'infezione, e il livello residuo di inoculo da parte di questa crittogama. Con il modello previsionale proposto da ERSA sarà possibile osservare le seguenti variabili descrittive dell'evoluzione della malattia:
- La dose di ascospore maturata giornalmente, nel periodo delle infezioni primarie (stagione in cui si concentra la formazione della quasi totalità delle spore primarie);
- Gli eventi di rilascio delle ascospore, individuati secondo specifiche situazioni condizionali;
- Gli eventi di dispersione delle ascospore e le dosi di ascospore depositate, determinate dalla pioggia e dalle temperature;
- Gli eventi di infezione primaria vera e propria, determinati dalla durata della bagnatura (ore) e dalle temperature;
- Infine, gli indici di rischio infettivo (Infection risks), calcolati sulla base della densità delle ascospore rilasciate e della suscettibilità dei tessuti ospiti, al fine della determinazione della gravità delle infezioni stesse.
Le informazioni simulatorie fornite dal modello forniranno le indicazioni di partenza per pianificare gli interventi fitosanitari in grado di prevenire il più possibile le infezioni primarie, in line con quanto previsto attualmente dalle strategie di produzione integrata.
Risultati elaborazione Ticchiolatura
Note sull'utilizzazione: risultati a carattere sperimentale in corso di validazione
Elaborazione modello
Fenologia del MELO su scala BBCH sulla base del calcolo dei gradi giorno cumulati (GDD)
| Fase vegetativa | |
| Fase fioritura | |
| Fase accrescimento frutto (solo arboree) | |
| Fase maturazione |
Carpocapsa (Cydia pomonella (Linnaeus, 1758))
 La carpocapsa è un lepidottero carpofago originario del centro Europa la cui diffusione, originariamente legata a quella del melo selvatico (sua pianta ospite iniziale) si è estesa in Asia, America del nord e sporadicamente nelle regioni dell'emisfero australe. La Carpocapsa è in grado di parassitare molte piante da frutto e rimane un insetto assolutamente attuale nelle strategie di difesa delle pomacee e del noce, ospiti principali del fitofago, ma può interessare anche i frutti di cotogno, nespolo e kaki e più raramente albicocco, susino e pesco.
Le larve di carpocapsa sono di colore bianco-roseo con capo scuro e si differenziano dalle larve di Cydia molesta (Tignola orientale del pesco), altro insetto carpofago che arreca danni al melo, per l'assenza del pettine anale.
Gli adulti hanno un'apertura alare di circa 20 mm, con ali anteriori di colore grigio scuro e linee ondulate trasversali. Nella parte posteriore delle ali è presente un disegno a forma di occhio di colore marrone scuro.
La carpocapsa è un lepidottero carpofago originario del centro Europa la cui diffusione, originariamente legata a quella del melo selvatico (sua pianta ospite iniziale) si è estesa in Asia, America del nord e sporadicamente nelle regioni dell'emisfero australe. La Carpocapsa è in grado di parassitare molte piante da frutto e rimane un insetto assolutamente attuale nelle strategie di difesa delle pomacee e del noce, ospiti principali del fitofago, ma può interessare anche i frutti di cotogno, nespolo e kaki e più raramente albicocco, susino e pesco.
Le larve di carpocapsa sono di colore bianco-roseo con capo scuro e si differenziano dalle larve di Cydia molesta (Tignola orientale del pesco), altro insetto carpofago che arreca danni al melo, per l'assenza del pettine anale.
Gli adulti hanno un'apertura alare di circa 20 mm, con ali anteriori di colore grigio scuro e linee ondulate trasversali. Nella parte posteriore delle ali è presente un disegno a forma di occhio di colore marrone scuro.
Ciclo biologico
Il ciclo dell'insetto è regolato dalla temperatura e dal fotoperiodo. In Friuli Venezia Giulia compie, di solito, due generazioni complete all'anno, ma non sono rare anche le tre generazioni a seconda degli stimoli esterni. Il suo sviluppo è condizionato dall'andamento climatico e, in coincidenza di alte temperature e bassa piovosità, le popolazioni aumentano risultando più aggressive.
Completato il ciclo il fitofago entra in uno stato di diapausa, tessendo il bozzolo sotto la corteccia delle piante o nell'anfrattuosità del tronco o dei rami più grossi, per attraversare la stagione autunno-invernale. Tale processo è influenzato da fattori in parte climatici (temperature, fotoperiodo) e in parte biologici (dieta delle larve di ultima generazione).
In primavera, circa due settimane prima della fuoriuscita dell'adulto, le larve si incrisalidano (marzo-aprile) e quando vengono raggiunti i 140-150 Gradi Giorno (GG*), si ha l'inizio del primo volo. Nel nostro areale questo avviene generalmente dalla prima decade di aprile per poi concludersi tra fine maggio e metà giugno. Gli adulti sono attivi al tramonto e con temperature crepuscolari superiori ai 13-15 °C avvengono gli accoppiamenti seguiti, dopo alcuni giorni, dalle ovideposizioni che avvengono inizialmente solo su foglie e successivamente anche sui frutti.
Le uova sono deposte in genere in modo isolato o al massimo accoppiate e leggermente sovrapposte. Le prime ovideposizione iniziano al raggiungimento dei 230 GG circa.
Il successivo sviluppo embrionale necessita di 90 GG per completarsi e, al raggiungimento dei 310 GG, avvengono le prime nascite larvali. Le larve neonate trascorrono alcuni giorni all'esterno vagabondando tra le foglie, che possono anche "danneggiare", per poi penetrare nei frutti. Generalmente ciò avviene al raggiungimento dei 325-330 GG. All'interno dei frutti le larve completano il loro sviluppo in 21-30 giorni circa e, una volta mature, fuoriescono dai frutti infestati per andare ad incrisalidarsi sotto la corteccia o altri ripari. Gli adulti di seconda generazione compaiono da fine giugno a tutto agosto e anche oltre (880 GG), con un picco massimo nella prima metà di luglio. Una parte delle larve di seconda generazione completa il ciclo iniziando un terzo volo a metà agosto, sovrapponendosi talvolta al secondo volo. Le larve dell'ultima generazione completano lo sviluppo nei frutti in maturazione per poi entrare in diapausa. Le penetrazioni delle larve di questa seconda generazione avvengono al raggiungimento dei 990-1000 GG circa.
* somma dei gradi di temperatura media che dall'inizio dell'anno superano i 10 °C
Sintomatologia e danni
La dannosità dell'insetto è legata al suo elevato potenziale biotico e alle notevoli capacità di perdite produttive che è in grado di causare. Le larve di carpocapsa infatti sono carpofaghe, ossia si nutrono direttamente del frutto, per questo la soglia di danno è molto bassa.
I danni arrecati alla produzione possono variare molto da un anno all'altro, anche all'interno della stessa azienda, e tra le diverse cultivar. Nel melo le varietà più suscettibili sono quelle del gruppo delle Red Delicious, mentre nel pero la cultivar più interessata sembra essere l'Abate Fetel.
L'organizzazione della difesa deve tener conto di queste variazioni e dovrà essere tarata di volta in volta nella propria realtà aziendale.
Nelle mele e pere, le larve di prima generazione entrano principalmente dalla zona calicina del frutto, ma anche attraverso un qualsiasi altro punto dell'epicarpo, soprattutto nei punti di contatto fra un frutto e l'altro. Creando delle gallerie nella polpa le larve raggiungono quindi i semi di cui si nutrono affinché la larva possa trasformarsi in adulto. Poco dopo la penetrazione nel frutto, in corrispondenza del foro di entrata, sono visibili dall'esterno delle "rosure" ed escrementi che consentono la rapida individuazione della sua presenza e del danno arrecato. Nei frutti immaturi la larva penetra nella zona sottoepidermica scavando una galleria a spirale per poi dirigersi verso la zona carpellare dove raggiunge i semi. Nei frutti maturi invece la larva entra direttamente nella polpa del frutto e raggiunge i semi per mangiare. Di solito non si trova più di una larva per frutto per il fenomeno del cannibalismo.
Monitoraggio
Effettuare il monitoraggio della carpocapsa con le trappole a feromoni è fondamentale per attuare le corrette strategie di difesa. Le trappole vanno posizionate in campo entro la metà aprile (attorno ai 100-120 GG).
Si precisa che il monitoraggio del solo volo con le trappole a feromoni non può mai essere indicativo dell'attività della carpocapsa. Rimangono pertanto fondamentali ed importanti i controlli visivi per valutare l'evoluzione e la situazione reale in campo. Tali verifiche devono interessare almeno 1000 frutti/ha o 500 frutti/appezzamento ad iniziare dal raggiungimento dei 330 GG (inizio delle prime penetrazioni e quindi dei danni visibili sui frutti) per la prima generazione e ai 1000 GG alla ripresa delle penetrazioni estive.
Strategie di difesa
La carpocapsa, vivendo come larva all'interno dei frutti, non ha molti nemici naturali. Tuttavia, si possono adottare alcuni accorgimenti che permettono di ridurre la popolazione dell'insetto quali:
- Posizionamento di nidi artificiali per uccelli insettivori che si nutrono degli adulti di carpocapsa e delle larve quando sono ancora gregarie all'esterno dei frutti;
- Favorire la presenza di insetti antagonisti naturali (es. forficula auricularia) ed imenotteri (Braconidi, Icneumonidi);
- Evitare condizioni favorevoli per il suo sviluppo/ripopolamento quali ad esempio la vicinanza ad impianti di noce, cumuli di legna, fonti luminose che attirano di sera glia adulti.
Per il contenimento delle popolazioni di carpocapsa è possibile adottare tecniche di confusione e distrazione sessuale, idonee per frutteti di conveniente ampiezza, andando a posizionare, prima dell'inizio dei voli dell'annata, gli specifici dispenser erogatori che, a seconda del tipo, possono bastare per l'intera stagione o che richiedono di essere rinnovati. L'applicazione dei diffusori deve terminare entro i 120 GG. Anche nei frutteti con confusione è importante posizionare le trappole a feromoni per verificare l'efficacia della confusione/disorientamento. Laddove non vengano adottate le tecniche di confusione o distrazione sessuale, le strategie di difesa consistono nel contenere il più possibile la prima generazione dell'insetto mediante trattamenti insetticidi nella fase di ovideposizione ed eventualmente con ulteriori interventi alla generazione larvale, avendo cura di bagnare sempre bene le piante.
La seconda generazione viene controllata alternando le molecole per evitare l'insorgenza di fenomeni di resistenza. In Friuli Venezia Giulia è possibile prendere visione dei principi attivi previsti dal Disciplinare di Produzione Integrata (DPI) per la difesa integrata volontaria.
Per un efficacie controllo di questo insetto risulta fondamentale il monitoraggio attraverso l'utilizzo delle trappole a feromoni per controllarne costantemente lo sviluppo fenologico. Ad esempio, studi sperimentali condotti in altre regioni e adottate anche nei DPI prevedono che la soglia di intervento sia fissata con la presenza di 2 adulti per trappola in 1 o 2 settimane per la I e II generazione. In caso di infestazioni posso essere prese in considerazione le soglie con le percentuali di danno sui frutti.
Modello previsionale
Il modello riportato consente, mediante funzioni che esprimono il tasso/la velocità di sviluppo, di individuare preventivamente, durante la stagione, l'andamento delle diverse fasi fenologiche, gli eventi di ovodeposizione e mortalità e il numero di stadi non osservabili. Questo consente di effettuare i monitoraggi e gli interventi di difesa nel momento più adatto.
Risultati elaborazione Carpocapsa
Note sull'utilizzazione: risultati a carattere sperimentale in corso di validazione
Curve fenologiche normalizzate
Fenologia del MELO su scala BBCH sulla base del calcolo dei gradi giorno cumulati (GDD)
| Fase vegetativa | |
| Fase fioritura | |
| Fase accrescimento frutto (solo arboree) | |
| Fase maturazione |
Diabrotica del mais (Diabrotica virgifera virgifera (Le Conte, 1868))
 La diabrotica del mais è un coleottero crisomelide di origine americana, diffusosi in Europa dell'est all'inizio degli anni '90, segnalato in Italia nel 1998 (provincia di Venezia) e successivamente diffusosi in tutto il nord del Paese.
La diabrotica compie una generazione all'anno ed il ciclo biologico è caratterizzato dagli stadi di uovo, larva, pupa e adulto; la comparsa e la durata dei diversi stadi dipende dalle condizioni ambientali stagionali.
Le uova, della dimensione di circa 0,5 mm, sono di colore giallo pallido. Le larve, di colore biancastro con testa scura, raggiungono a maturità una lunghezza variabile fra 10 e 18 mm e le pupe sono di colore bianco. Gli adulti presentano una lunghezza variabile tra 4 e 6,5 mm. I maschi hanno in genere un colore più scuro delle femmine e sono dotati di elitre scure con due macchie gialle all'estremità dell'addome. Le femmine invece presentano due striature gialle su tutta la lunghezza delle ali. La lunghezza delle antenne è maggiore nei maschi nei quali raggiunge o supera quella del corpo, mentre nelle femmine non supera i 3/4 della lunghezza corporea.
La diabrotica del mais è un coleottero crisomelide di origine americana, diffusosi in Europa dell'est all'inizio degli anni '90, segnalato in Italia nel 1998 (provincia di Venezia) e successivamente diffusosi in tutto il nord del Paese.
La diabrotica compie una generazione all'anno ed il ciclo biologico è caratterizzato dagli stadi di uovo, larva, pupa e adulto; la comparsa e la durata dei diversi stadi dipende dalle condizioni ambientali stagionali.
Le uova, della dimensione di circa 0,5 mm, sono di colore giallo pallido. Le larve, di colore biancastro con testa scura, raggiungono a maturità una lunghezza variabile fra 10 e 18 mm e le pupe sono di colore bianco. Gli adulti presentano una lunghezza variabile tra 4 e 6,5 mm. I maschi hanno in genere un colore più scuro delle femmine e sono dotati di elitre scure con due macchie gialle all'estremità dell'addome. Le femmine invece presentano due striature gialle su tutta la lunghezza delle ali. La lunghezza delle antenne è maggiore nei maschi nei quali raggiunge o supera quella del corpo, mentre nelle femmine non supera i 3/4 della lunghezza corporea.
Ciclo biologico
La diabrotica compie una generazione l'anno: lo svernamento avviene allo stadio di uovo. Le uova vengono deposte nella parte superficiale del terreno, nelle porzioni più umide e la loro schiusura inizia a metà maggio, con un picco delle nascite in giugno. Le larve vivono nel terreno e, prima di impuparsi, compiono tre stadi di sviluppo in un periodo di tempo variabile da 30 a 50 giorni a seconda della temperatura. Esse si muovono nel terreno nutrendosi in particolare delle radici del mais ed anche di altre specie erbacee. I primi danni alle radici avvengono subito dopo la loro comparsa ma le conseguenze sono particolarmente evidenti tra la fine di giugno ed i primi di luglio. La durata dello stadio pupale è di 1-2 giorni a 22 °C di temperatura. L'inizio dei voli avviene nella seconda metà di giugno con lievi variazioni in base all'andamento stagionale, e continua fino ad ottobre. La presenza degli adulti raggiunge la maggiore consistenza in luglio (nel periodo della fioritura del mais) e prosegue poi fino ad autunno inoltrato. La temperatura ottimale per lo sviluppo degli adulti è compresa tra 23 e 27 °C. Nelle ore più calde, quando la temperatura supera i 30 °C gli insetti si rifugiano in gruppi sulle parti della pianta meno esposte al sole. Gli accoppiamenti iniziano circa una settimana dopo l'inizio dei voli, sia i maschi che le femmine possono accoppiarsi più volte nel corso della loro vita. Gli adulti, oltre che a spostarsi dal basso verso l'alto sulla singola pianta e tra le piante dello stesso campo, spesso vengono trasportati dal vento o da altri vettori (tra cui l'uomo) e si diffondono anche da un campo all'altro, a volte riuscendo a coprire grandi distanze. Lo sviluppo di D. virgifera e l'incremento delle sue popolazioni è favorito dalla coltivazione di mais in monosuccessione, in particolare su terreni con buone proprietà chimico-fisiche e biologiche, ricchi di sostanza organica, con PH da sub acido a sub alcalino e con buona disponibilità idrica. Il tasso di mortalità di uova e larve è maggiore nei terreni compatti e pesanti. Le condizioni meteorologiche influenzano le popolazioni dell'insetto: inverni miti e primavere umide facilitano la sopravvivenza delle uova, come le estati fresche, che favoriscono l'attività degli adulti. La mortalità durante la diapausa invernale è elevata solo in presenza di inverni particolarmente rigidi. Le larve prediligono condizioni di buona umidità, ma sono estremamente sensibili agli eccessi idrici che sono un importante fattore di mortalità durante la schiusa delle uova e la comparsa delle larve di prima età.
Sintomatologia e danni
I danni causati dall'insetto alle colture di mais comprendono l'allettamento delle piante a causa dell'attacco delle larve all'apparato radicale e l'erosione delle infiorescenze e delle parti epigee a seguito dell'attività trofica degli adulti. Il danno alle radici, che è quello più importante e consistente, viene procurato dalle forme larvali. Per convenzione viene classificato secondo una scala di crescente gravità (scala Iowa), che va da un leggero attacco alle radici più esterne, che procura solo qualche cicatrice ed è irrilevante dal punto di vista economico (grado 1), fino alla distruzione dell'intero apparato radicale (grado 6). In quest'ultimo caso si assiste ad una diminuzione consistente del raccolto che rimane a terra per allettamento delle piante. Nei casi di elevata infestazione, ad aggravare il danno larvale si aggiunge l'attività trofica degli adulti nei confronti delle parti epigee con particolare predilezione per le sete fiorali, limitando anche gravemente l'allegagione; dopo la fioritura vengono attaccate anche le spighe con cariossidi allo stadio ceroso.
Monitoraggio
L'attività di monitoraggio inizia dalla prima settimana di maggio con l'osservazione diretta in campagna e con il campionamento del terreno per l'individuazione delle larve, prosegue con il controllo settimanale delle trappole munite di feromone sessuale (n.3 trappole per appezzamento), per l'individuazione del primo volo dell'insetto adulto, e delle trappole cromotropiche per il monitoraggio dell'andamento dello sviluppo degli individui adulti (almeno n.2 trappole per appezzamento). Un metodo semplice per controllare la presenza di larve in campo è quello di prelevare una zolla di terreno dall'intorno di una radice e sgretolarla su un sacco di plastica nero, in questo modo le larve bianche si notano facilmente per contrasto.
Il monitoraggio degli insetti consente di valutare l'opportunità di intervenire con appositi trattamenti adulticidi quando la loro presenza diviene significativa (trattamento che non va eseguito se la coltura si trova in fase di fioritura), per controllare le ovideposizioni e valutare il rischio di danno per l'anno successivo, qualora si seminasse nuovamente il mais.
Strategie di difesa
Alla luce del fatto che il rischio di danno si presenta sugli appezzamenti a mais in monosuccessione, per più anni e solamente in aree dove la densità di popolazione è elevata, lo strumento più efficace di contrasto e lotta alla diabrotica consiste nell'applicazione di tecniche agronomiche come la rotazione colturale.
Nel caso in cui si possa ricorrere ad una semina ritardata, questa risulta essere molto efficace nel caso in cui venga effettuata dopo la comparsa delle larve che, non trovando nutrimento sufficiente al momento della nascita, muoiono di fame.
Tecniche colturali che favoriscono lo sviluppo dell'apparato radicale (es. concimazione, rincalzatura, ecc.) influenzano positivamente lo sviluppo della pianta nonostante la presenza dell'insetto. Nella scelta dell'ibrido, sono da preferire quelli precoci poiché consentono alla pianta di giungere con le radici maggiormente sviluppate nel periodo di massima presenza dell'insetto. Sono inoltre da privilegiare ibridi con apparato radicale particolarmente vigoroso e con una buona resistenza all'allettamento. L'utilizzo di geodisinfestanti produce buoni risultati se questi vengono utilizzati all'approssimarsi della comparsa delle prime larve, tenendo comunque in considerazione i fattori che possono influire sulla loro efficacia (andamento pluviometrico, tipologia di terreno, sostanza attiva, etc.).
È possibile effettuare anche un trattamento adulticida che fornisce i migliori risultati se effettuato prima che le femmine abbiano deposto un numero significativo di uova. È necessario quindi controllare il livello di popolazione in campo negli appezzamenti in cui si intende seminare mais anche l'anno successivo. Secondo le direttive della lotta integrata il trattamento va eseguito solamente nel caso in cui si rileva una media di individui superiore a 50 insetti per trappola cromotropica durante due settimane consecutive. In ogni caso, il trattamento non deve essere eseguito se il mais si trova in fase di fioritura.
Modello previsionale
Il modello previsionale proposto consente di prevedere anticipatamente lo sviluppo fenologico dell'insetto, attraverso la visione di curve fenologiche e di curve di emergenza degli adulti, nonché i gradi giorno cumulati per lo stadio pre-adulto e adulto. Il momento in cui la curva dei gradi giorno raggiunge la massima pendenza è quello in cui si ha il picco di presenza dell'insetto in un determinato stadio e in base ad esso si possono valutare le eventuali misure di contenimento da adottare.
Risultati elaborazione Diabrotica
Note sull'utilizzazione: risultati a carattere sperimentale in corso di validazione
Curve di emergenza degli stadi pre-adulto
Curve fenologiche di larve, pupe e adulti
Gradi giorno cumulati
Fenologia del MAIS su scala BBCH sulla base del calcolo dei gradi giorno cumulati (GDD)
| Fase vegetativa | |
| Fase fioritura | |
| Fase maturazione |
Piralide del mais (Ostrinia nubilalis (Hübner, 1796))
La Piralide del mais (Ostrinia nubilalis Hübner, 1796) è un insetto appartenente alla famiglia Crambidae, dell'ordine dei Lepidotteri, il cui sviluppo si suddivide nelle quattro fasi fenologiche di uovo, larva, pupa e adulto. È un insetto polifago, le sue larve sono in grado di danneggiare un notevole numero di piante erbacee, sia coltivate che spontanee (peperone, fagiolino, barbabietola, canapa, ecc.) ma il mais resta comunque il principale bersaglio di questo insetto.
La larva misura circa 20-25 mm di lunghezza, presenta una livrea di colore grigiastro o nocciola con bande longitudinali, una mediana e due laterali, tendenti al verde con file di tubercoli scuri. Il capo e il protorace sono bruno scuro. L'adulto è caratterizzato da dimorfismo sessuale: la femmina ha un'apertura alare di circa 30 mm, con ali colore crema con linee scure dentellate. Il maschio misura circa 25 mm, con ali anteriori giallastre, con variegature più scure.
Ciclo biologico
L'insetto sverna all'interno delle parti di piante morte lasciate sul campo, a livello di quinto stadio larvale, compiendo la diapausa. In Italia settentrionale la Piralide compie due o tre cicli all'anno: lo sfarfallamento dei lepidotteri svernanti avviene verso fine maggio, seguito dall'accoppiamento e dalla conseguente ovideposizione. Il secondo volo, che dà origine alla seconda generazione di larve, raggiunge il picco tra fine luglio e agosto, a seconda delle condizioni climatiche del momento. Può verificarsi anche un terzo volo nel mese di settembre che a sua volta genera la terza generazione di larve.
Sintomatologia e danni
Le larve di prima generazione generalmente non recano grossi danni dal momento che, una volta fuoriuscite dall'uovo, si nutrono delle parti verdi della pianta di mais attaccando le guaine fogliari, le foglie ed entrando nel culmo, dove, raggiunta la maturità, si incrisalidano. Quelle di seconda generazione invece agiscono sulla spiga e sulle cariossidi, arrecando maggior danno sia in termini di produzione (resa e qualità del prodotto) sia in termini di tossicità, in quanto all'interno dei fori e delle gallerie scavate si possono insediare funghi parassiti (Aspergillus, Fusarium) che possono sintetizzare micotossine cancerogene.
Ricapitolando, i danni sono attribuibili alla piralide sono essenzialmente 3: fisiologici, qualitativi e quantitativi. I danni fisiologici comprendono la formazione di gallerie che porta ad una alterazione delle funzionalità metabolica della pianta impedendo in tal modo il normale flusso di acqua, sostanze nutritive ed elementi prodotti dalla fotosintesi. Questo causa un decremento produttivo proporzionale all'attacco dell'insetto. Quelli quantitativi sono legati allo stroncamento dei tutoli, sempre a causa delle gallerie presenti, e alla perdita della pannocchia durante la fase di raccolta. Infine, per quanto riguarda i danni qualitativi, le gallerie e i fori causati dall'attacco dell'insetto costituiscono la via di accesso preferenziale di alcuni funghi parassiti. Le principali conseguenze che si registrano sono: mancato prodotto durante la raccolta, spighe e cariossidi danneggiate, difficoltà di stoccaggio e d'insilaggio e non per ultimo per importanza disturbi digestivi e di tossicità nei confronti degli animali. La diretta e importante conseguenza è una minore produzione e una qualità scadente della granella. Il risvolto economico riguarderà il mancato guadagno per la quantità di mais non raccolto, il mancato guadagno del mais non conservabile che non verrà così utilizzato e, gli aspetti qualitativi di ordine igienico – sanitario, che per loro natura sono difficilmente quantificabili.
Monitoraggio
L'attività di monitoraggio inizia verso la metà di maggio attraverso la cattura degli adulti con trappole a cono di rete munite di feromoni, oppure con trappole luminose al neon, posizionate sul bordo dell'appezzamento coltivato a mais, e consiste nel conteggio bisettimanale degli insetti adulti catturati. In questo modo viene monitorato sia l'andamento che la consistenza dei voli, al fine di valutare l'opportunità di intervenire con appositi trattamenti.
Strategie di difesa
La lotta alla Piralide può essere condotta attraverso mezzi agronomici, chimici, biologici/biotecnologici. I primi consistono nella rotazione colturale, seguita dalla distruzione dei residui di mais (sfibratura degli stocchi ed aratura). Al fine di eliminare le larve svernanti, è importante inoltre la corretta scelta delle varietà, l'anticipo della semina, l'utilizzo di classi varietali a ciclo breve, l'impiego di cultivar caratterizzate da uno stocco robusto e l'esecuzione di irrigazioni a pioggia successive alla fioritura.
In caso di forti attacchi si possono effettuare degli interventi chimici alla comparsa della prima generazione e trattamenti sulla seconda generazione qualora vi sia la necessità. I trattamenti devono essere eseguiti in funzione dei voli rilevati con le trappole ma non durante la fase di fioritura. È possibile effettuare sia trattamenti chimici che biologici. Nell'applicazione dei prodotti e nella scelta della tempistica è necessario prendere in considerazione la scalarità delle ovideposizioni, la natura endofita del lepidottero e la variabilità ambientale. La piralide, infatti, segue le proprie dinamiche biologiche con ritmi e tempi diversi nelle singole annate, evolvendo talora indipendentemente dallo stadio di sviluppo del mais.
Per quanto riguarda la lotta biologica, si possono utilizzare preparati a base di Bacillus thuringiensis (batterio sporigeno che vive nel terreno), oppure attraverso l'utilizzo dell'Imenottero Trichogramma maidis al fine di assicurare un alto grado di parassitizzazione delle uova della Piralide.
Modello previsionale
Il modello previsionale consente di identificare le fasi fenologiche dell'insetto con i rispettivi picchi di presenza attraverso il calcolo dei gradi giorno. In base a questi è possibile individuare il momento migliore per mettere in atto le strategie di difesa descritte.
Risultati elaborazione Piralide
Note sull'utilizzazione: risultati a carattere sperimentale in corso di validazione
Curve fenologiche
Gradi giorno cumulati
Fenologia del MAIS su scala BBCH sulla base del calcolo dei gradi giorno cumulati (GDD)
| Fase vegetativa | |
| Fase fioritura | |
| Fase maturazione |